FISH techniques
Table of contents
1.1.1. INTRODUCTION
1.1.2. FISH VARIANTS
1.1.3. FISH APPLICATIONS IN MEDICINE
1.1.4. SOURCES:
FISH techniques [edit]
(Emanuela Boccuni)
INTRODUCTION
Fluorescence in situ hybridization (FISH) is a nonradioactive cytogenetic technique which uses fluorescent probes binding only those parts of the chromosome with a high degree of sequence complementarity. It was developed in 1980 and had the goal to identify, and eventually quantify, the presence of given DNA sequences on chromosomes in order to define a spatial-temporal pattern of gene expression. FISH can be also used to detect and localize specific RNA targets, coding and non-coding RNA in cells, tumours and tissue samples.
The possibility to identify the exact positioning of a DNA sequence is the basic concept of in situ approaches. Historically, FISH and other in situ hybridization results played a primary role in mapping genes on human chromosomes. Results from these experiments were collected in databases, and they became very useful for the progress of the Human Genome Project (HGP).
From its development in the 1960s onwards, FISH largely benefitted from improvements of probe-labeling techniques and specific probe design strategies increasing its sensitivity. Its broad application in research and diagnostics are shown by the rise of the number of publications reporting FISH at the beginning of the early 1990s. In recent years, the combination of microfluidic techniques and FISH addresses limitations in probe consumption and hybridization times, making the experimental procedure more sustainable and adaptable to high-throughput developments. Now that the HGP is complete, scientists rarely use in situ hybridization simply to identify the chromosomal location of a human gene: in fact, nowadays there are many applications of this technique, principally focused on clinical diagnoses.
In the figure below, a timeline of fluorescence in situ hybridization developments.
The earliest record of in situ hybridization is found by Gall and Pardue in 1969. First fluorescent versions of the technique (FISH) appeared in the 1970s, followed by direct probe labeling twenty years later. ‘Modern’ FISH includes developments in the probe design and production. The combination of microfluidics and FISH first appeared in the early 21st century. Events directly related to the development of FISH are shown in boxes.
In this technique, DNA or RNA probes detect segments of the human genome by DNA-DNA hybridization,DNA sample in a metaphase stage previously treated under conditions that preserve the morphology of condensed human chromosome. Probes are labelled at the 3’ end, usually, with fluorescein or other fluorochromes, that have been replaced the radioactive labels because of their greater safety.
Probes can be often derived from an entire sequence of DNA, which has been cut by sequence-specific endonucleases, isolated, purified, and amplified, firstly for use in the Human Genome Project and later for subsequent studies in the same field.
Results are detected by a fluorescence microscopy, used to find out where the fluorescent probe bound to the chromosome: the hybridization reaction generates the fluorescent light.
It is a powerful tool for understanding a variety of chromosomal abnormalities and other genetic mutations. FISH differs from the other techniques for the possibility to perform the procedure both in divided and non-divided cells, making it a very versatile technology.
Moreover, FISH is used for examining the cellular reproduction cycle, specifically interphase of the nuclei for any chromosomal abnormalities.

(Marica Zoppi)
FISH VARIANTS
FISH, like other DNA-based approaches, relies on the ability of nucleic acids to de- and renature and of homologous sequences to find each other and build a double helix again.
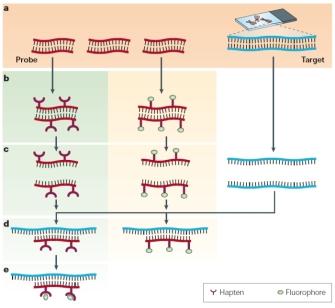
The first step is to prepare a DNA probe specific for the the gene of interest. The DNA probe can be labeled in various ways, such as nick translation, random primed labeling, and PCR. There are two labeling strategies:
direct labeling, meaning that the fluorochromes are directly bound to the probe DNA
indirect labeling, where a non-fluorescent hapten bind to the fluorophore subsequently
The probe and target sequences are then mixed together , and the probe specifically hybridizes to its complementary sequence on the chromosome. If the probe is already fluorescent (middle column), it will be possible to detect the site of hybridization directly. In other cases (left column), an additional step may be needed to visualize the hybridized probe. Hybrids formed between the probes and their chromosomal targets can be detected using a fluorescent microscope.
Then, the labeled probe and the target DNA are denatured with heat or chemicals, this denaturation step is important for the formation of new hydrogen bonds between the target and the probe. The probe and target sequences are mixed together and the probe specifically hybridizes to its complementary sequence on the chromosome.
If the probe is already fluorescent (direct labelling), it will be possible to detect the site of hybridization directly. In case of a indirect labelling an additional step is required to visualize the hybridized probe. A fluorescent microscope detect the hybrids formed between the probes and their chromosomal targets.
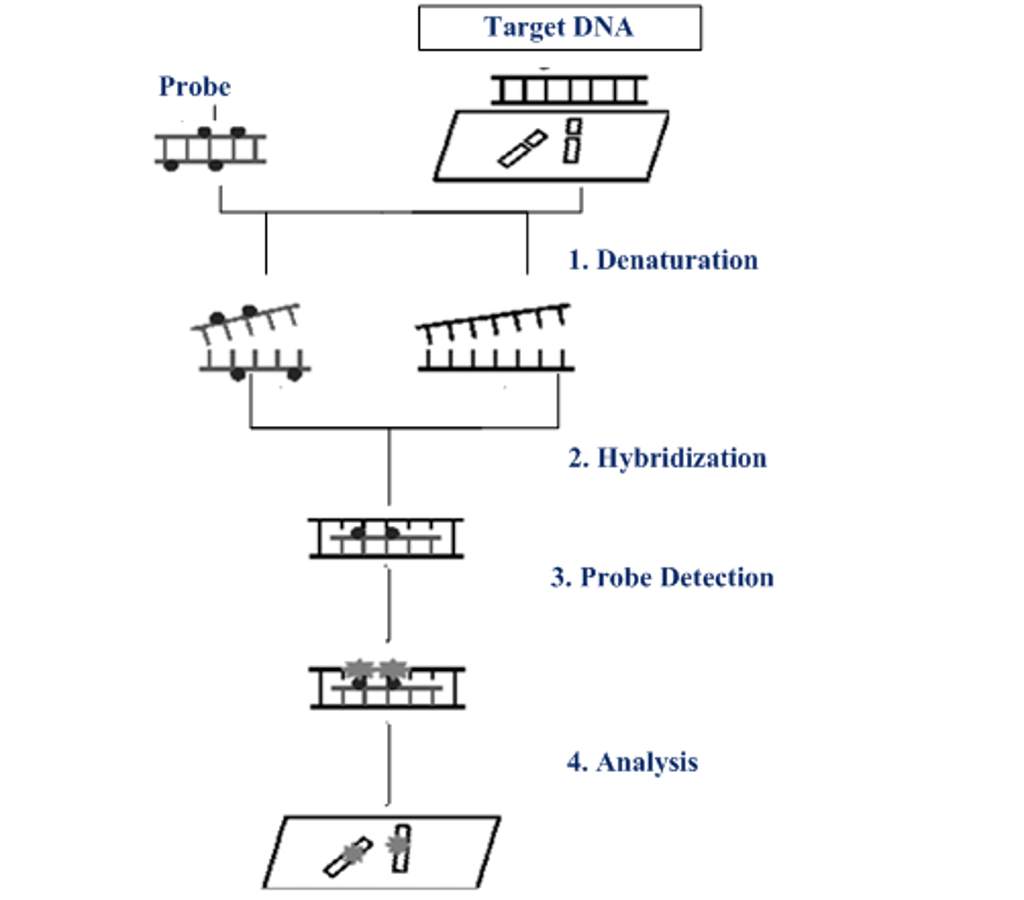
FISH is a less time-consuming technique in comparison to the conventional method of cytogenetic metaphase karyotype analysis. It is useful to detect specific cytogenetic abnormalities, as well as a copy of aberrations numbers. FISH is able to spot chromosomal microdeletion, amplification and translocation, for this reason is very useful diagnostic tool. Many different types of FISH are now available to cytogeneticists and its sensitivity has increased enormously.
MULTIPLEX FISH or M-FISH
Multiplex in situ hybridization (M-FISH) is a 24-color karyotyping technique used for studying complex interchromosomal rearrangements.
The process involves three major steps.
- Firstly, each homologous is labelled with finite numbers of spectrally distinct fluorophores, such that each homologous pair of chromosomes is uniquely labeled. A multiplex probe set for M-FISH consists of whole chromosome probes (WCP) each labeled with one of spectrally distinct fluorophores. The majority of commercially available human probe sets consist of 52 WCP directly and indirectly labeled with one of five fluorophores, in this way each chromosome is uniquely labeled entirely down its whole length with one, two, or three different fluors.This type of labeling is called combinatorial and it is at the base of M-FISH chromosome classification.
- Secondly, the microscopic visualization and digital acquisition of each fluorophore using a specific software for M-FISH. The process of M-FISH image acquisition for each metaphase spread involves the digital capture of five fluorescence channels and the DAPI counterstain in sequence. Once acquired the digital image, it’s possible to classify the chromosomes according to the combinatorial labeling scheme of the M-FISH probe cocktail used.
- Finally, it’s possible to resolve structural and numerical abnormalities. In fact, the power of M-FISH (and SKY) lies in its ability to resolve complex karyotypes and identify aberrations.
M-FISH is a very powerful technique for detecting interchromosomal alterations of varying complexity, throughout the genome. However, it has some limitation since it cannot detect inversions, amplifications, deletions, or homologous pair rearrangements.
For standard M-FISH based on five fluorochromes and one counterstain, at the very least, a fluorescence microscope suitable for six filter sets is required. A charge-coupled device (CCD) camera connected to computer-based image acquisition and evaluation software is also needed. Overall therefore, a significant investment in hardware and software is necessary to build a good quality M-FISH system.
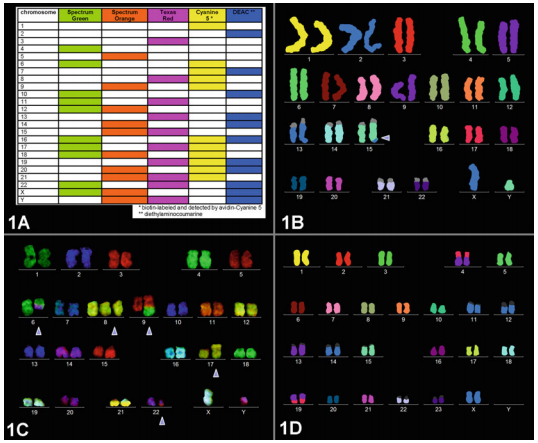
Fig. 1 (a) M-FISH labeling scheme for M-FISH set. (b) Prenatal case with a karyotype 46,XY,der(15)t(X;15)(q25;q26.3). The derivative chromosome 15 is denoted by an arrowhead. Here the M-FISH result is depicted in pseudo-colors. The heterochromatic regions unlabeled by the M-FISH probe set are shown in gray pseudo-colors.
(Maria Vittoria Di Marco)
Spectral Karyotyping (SKY)
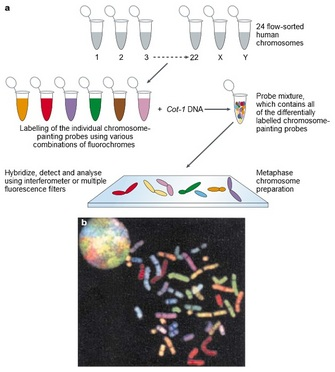
Spectral karyotyping (SKY) is a cytogenetic technique developed to display and identify the aberrations that can’t be detected by conventional banding technique on all 24 humans chromosomes at one time. By the simultaneous hybridization of multiple chromosome painting probes, each tagged with a specific fluorochrome or fluorochrome combination, it allows the visualization of individual chromosomes in metaphase ells and the identification of both numerical and structural chromosomal aberrations in human pathology with high sensitivity and specificity.
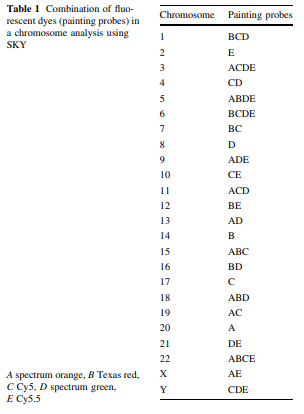
The color emission of chromosomes is determined by the combination of painting probes and fluorescent dyes; new colors can be developed by extracting a pair of different fluorescent dyes from among the five types of fluorescent dyes (spectrum orange, Texas red, Cy5, spectrum green, Cy5.5) and mixing each pair together. Theoretically 31 types of colors can be created from five types of fluorescent dyes; in reality however it is only possible to use 24 different distinguishable colors because some fluorescence has a wavelength similar to others within the infrared spectrum.
The complete procedure for SKY involves several steps and usually requires about one week time:
- metaphase cells’ slides are prepared after treatment with colcemid
- slides are treated with pepsin to remove the cytoplasm
- samples are incubated together with probes and they are both denatured to allow hybridization
- samples are analyzed: hybridization signals are captured using a fluorescent microscope equipped with a special CCD camera and image processing software.Color and inverted DAPI images of individual chromosomes are shown before the chromosomal sorting; after sorting, chromosomes are presented in the "classification" table.
This technique attributes a different color tones at each chromosome, making it possible to identify:
- chromosome rearrangements involving the whole or a portion of a chromosome(s)
- translocations involving two or more chromosomes
- marker chromosomes not resolvable by G-band karyotype analysis
A total of 20–50 cells are examined to interpret the overall chromosomal abnormalities: two or more cells with the same pattern of chromosomal aberration are considered a clonal abnormality.
SKY provides the advantage of easy visual interpretation when analyzing results, but it is not possible to evaluate structural abnormalities, such as inversion, deletion, insertion, and duplication in the same chromosome, because they are shown with the same color. In addition, the resolution limit of detection of approximately 1–2 Mb makes it impossible to detect smaller abnormalities. Therefore, in order to verify the site of chromosomal breakage, it is usually necessary to use SKY beforehand in combination with G-banding or high-resolution chromosome banding, instead of using SKY alone to macroscopically observe the banding patterns.
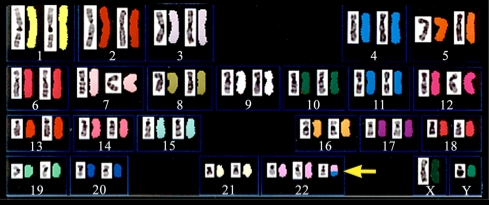
Fig. 1 - Chromosome analysis on Trisomy 11/22 using SKY technique.
3D FISH
Fluorescence in situ hybridization on three-dimensionally preserved nuclei (3D-FISH) in combination with 3D-microscopy and image reconstruction allows visualization of chromosome territories (CT), chromosome subregions, single genes, and RNA transcripts preserving their spatial positions in the cell nucleus. In many cases it is used in combination with immunocytochemistry to analyze the functional organization of the interphase nucleus, mapping DNA/RNA and protein targets in the same cell.
However, this combinatorial procedure has to be performed extremely carefully because some steps of the FISH protocol may interfere with immunostaining, making it unsuccessful.
In comparison with FISH on metaphase chromosomes, FISH on 3D-preserved nuclei requires special demands with regard to probe quality, fixation, and pretreatment steps of cells in order to achieve two goals: namely the best possible preservation of the nuclear structure and at the same time an efficient nuclear DNA accessibility for probes.
Probe labeling, hybridization, and post-hybridization steps are basically the same as for FISH on metaphase chromosomes:
- DNA probes are generated via different possible methods and are coupled with a fluorescent dye
- cells of interest are fixed to slides and permeabilized
- cellular DNA and directly labeled probes are then denatured together and hybridized overnight in a light-tight humidified chamber. The time depends on the number of layers used for detection. The choice of the detection scheme depends on several factors: the number fluorochromes used for probe labeling, the color of nuclear counter stain and the microscopic setup available.
- samples are washed, counterstained with DAPI and coverslips are mounted. An appropriate counterstaining is always necessary to test for preservation of normal nuclear morphology after FISH and to select well-preserved nuclei for the further analysis.
- an appropriate microscopy method (preferably confocal microscopy) is required for the analysis. A DNA peak at 260 nm is observed with a second peak corresponding to the absorbance wavelength of the fluorophore the probe is labeled with: probes with good incorporation have absorbance peaks of similar height or higher than the DNA peaks. Probes with a more than 2-fold higher fluorophore peak than DNA typically contain unconjugated dye that may cause background in the analysis.

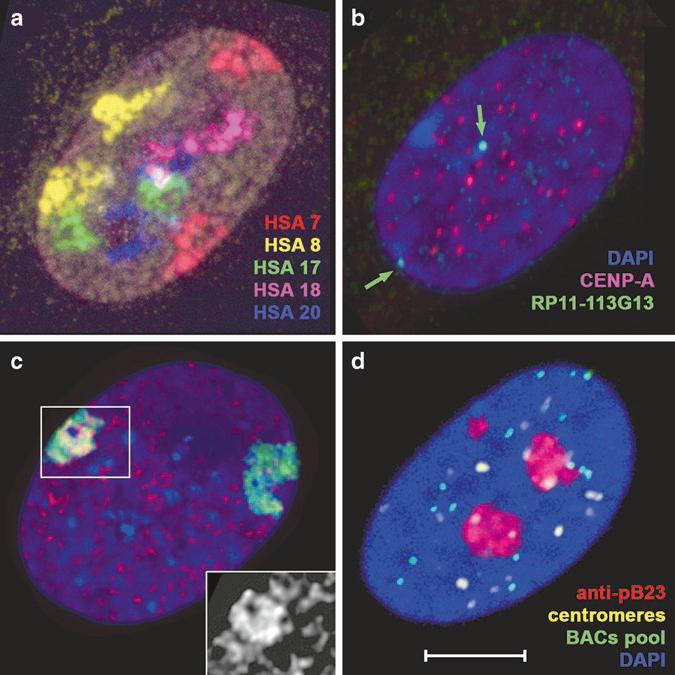
Fig.2 Examples of 3D-FISH and 3D-FISH combined with immunostaining. (a) Five-color chromosome painting. (b) Immunostaining before FISH. (c) Immunostaining before and after FISH (d) Immunostaining after FISH.
(Martina Ansaldi)
FISH APPLICATIONS IN MEDICINE
FISH can have many important applications in medicine; in particular it can be useful in:
- Identification of genetic aberrations: in general, aberrations can happen because of DNA breaks that can not be repaired by cellular mechanisms. Normally, from accumulation of these mutations, cancer can arise in human, and, nowadays, abnormalities in about 350 genes have been discovered as implicated in human cancers. Somatic mutations cancer genomes carry two types of somatic mutations: “driver” ones, as a result of selective pressure during tumorigenesis and positively selective, and “passenger” mutations, happening incidentally and results possible products of genome instability or large number of cell divisions. These last mutations normally lead to transformation of single cell in a detectable cancer.
These aberrations can be detected by FISH and can be therefore used to predict the development of cancer in a specific patient.
It can also be applied in the study of aberrations in prenatal and postnatal patients in order to predict the possible development of a specific disease.
- Detection of aberrations in many different chromosomes at the same time: SKY is an improvement of FISH application based on the use of fluorescent probes of different colors in order to visualize many different chromosomes together and distinguish them. This is important in the area of Gene Mapping.
- Another improvement of application in Gene Mapping is the ability to identify several different regions on the same chromosome by using, also in this case, different probes together. The process is called MULTIPLEX FISH and allow painting of the entire chromosome complement in a single hybridization through labelling each chromosome with a different combination of fluorophores.
- Comparative genomic hybridization to analyze chromosomal imbalance in tumors and to examine possible correlations between findings and tumor phenotypes.
- Study of chromosomal aberrations also in non dividing cells, leading to chromosomal mapping of some deleted or amplified region.
- Visualization and measurement of telomere length.
- Quantification of gene copy number and the amount of protein.
In general we can conclude saying that FISH in medicine finds its best applications in the correlation between DNA sequences (mutations and modifications inside it) and phenotype, usually cancer or disease. By using this process it can also be possible to predict the onset of a disease in a patient in different stages of his life.
SOURCES:
- “Application of Fluorescence In Situ Hybridization (FISH) Technique for the Detection of Genetic Aberration in Medical Science” 10.7759/cureus.1325
- “Applications of fluorescence in situ hybridization (FISH) in detecting genetic aberrations of medical significance” https://doi.org/10.1093/biohorizons/hzq009
- O'Connor, C. (2008) Fluorescence in situ hybridization (FISH). Nature Education 1(1):171
- “Chromosome Analysis Using Spectral Karyotyping (SKY)” 10.1007/s12013-011-9285-2